| |
Canku Ota
|
|
|
(Many Paths)
|
||
|
An Online Newsletter
Celebrating Native America
|
||
|
November
1, 2011 - Volume 9 Number 11
|
||
|
|
||
|
Western Honey Bee
|
||
|
From Wikipedia, the
free encyclopedia
|
||
|
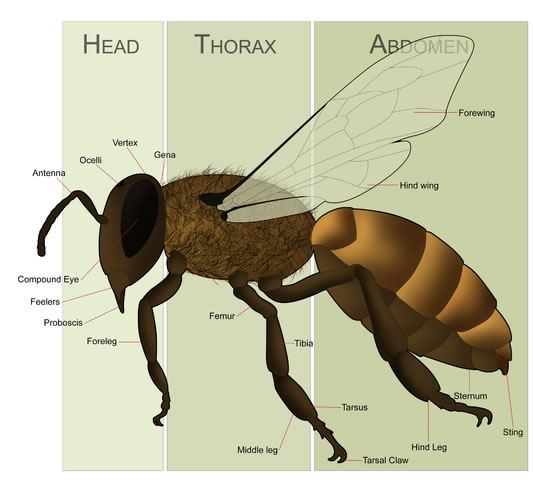
The Western honey bee or European honey bee (Apis mellifera) is a species of honey bee. The genus Apis is Latin for "bee", and mellifera comes from Latin melli- "honey" and ferre "to bear"—hence the scientific name means "honey-bearing bee". The name was coined in 1758 by Carolus Linnaeus who, realizing that the bees do not bear honey, but nectar, tried later to correct it to Apis mellifica ("honey-making bee") in a subsequent publication. However, according to the rules of synonymy in zoological nomenclature, the older name has precedence. As of October 28, 2006, the Honey Bee Genome Sequencing Consortium fully sequenced and analyzed the genome of Apis mellifera. In
2007 media attention focused on colony collapse disorder, a decline
in European honey bee colonies in a minority of regions of North
America.
Biology,
Life Cycle The larvae and pupae in a frame of honeycomb are referred to as frames of brood and are often sold (with adhering bees) by beekeepers to other beekeepers to start new beehives.
Both
workers and queens are fed "royal jelly" during the first
three days of the larval stage. Then workers are switched to a diet
of pollen and nectar or diluted honey, while those intended for
queens will continue to receive royal jelly. This causes the larva
to develop to the pupa stage more quickly, while being also larger
and fully developed sexually. Queen breeders consider good nutrition
during the larval stage to be of critical importance to the quality
of the queens raised, good genetics and sufficient number of matings
also being factors. During the larval and pupal stages, various
parasites can attack the pupa/larva and destroy or damage it. Worker
bees are infertile females, but in some circumstances, generally
only in times of severe stress or with the loss or injury or declining
health of the queen, they may lay infertile eggs, and in some subspecies
these eggs may actually be fertile. However, since the worker bees
are 'imperfect' females (not fully sexually developed), they do
not mate with drones. Any fertile eggs that they lay would be haploid,
having only the genetic contribution of their mother, and in honey
bees these haploid eggs will always develop into drones. Worker
bees also secrete the wax used to build the hive, clean and maintain
the hive, raise the young, guard the hive and forage for nectar
and pollen. Drones Life
Expectancy The lifespan of the workers varies drastically over the year in places with an extended winter. Workers born in the spring and summer will work hard and live only a few weeks, whereas those born in the autumn will stay inside for several months as the colony clusters. On average during the year about one percent of a colony's worker bees die naturally per day. Except for the queen, all of the colony's workers are therefore exchanged about every four months. Honey
Production When a hive detects smoke, many bees become remarkably non-aggressive; it is speculated that this is a defense mechanism. Wild colonies generally live in hollow trees, and when bees detect smoke it is presumed that they prepare to evacuate from a forest fire, carrying as much food reserve as they can. In order to do this, they will go to the nearest honey storage cells and gorge on honey. In this state they are quite docile since defense from predation is less important than saving as much food as possible. Thermoregulation Honey bees are able to forage over a 30 °C (54.0 °F) range of air temperature, largely because they have behavioural and physiological mechanisms for regulating the temperature of their flight muscles. From very low to very high air temperatures, the successive mechanisms are; shivering before flight and stopping flight for additional shivering, passive body temperature regulation in a comfort range that is a function of work effort, and finally, active heat dissipation by evaporative cooling from regurgitated honey sac contents. The body temperatures maintained differ depending on caste and expected foraging rewards.[4] The optimal air temperature for foraging is 22–25 °C (72–77 °F). During flight, the rather large flight muscles create heat, which must dissipate. The honey bee uses a form of evaporative cooling to release heat through its mouth. Under hot conditions, heat from the thorax is dissipated through the head; the bee regurgitates a droplet of hot internal fluid — a "honeycrop droplet" – which immediately cools the head temperature by 10 °C (18.0 °F).[5] Below 7–10 °C (45–50 °F) bees become immobile and above 38 °C (100 °F) bee activity slows. Honey bees can tolerate temperatures up to 50 °C (122 °F) for short periods. Queens
Regardless of the trigger, the workers develop the larvae into queens by continuing to feed them royal jelly which triggers an extended development as a pupa. When
the virgin queen emerges, she is commonly thought to seek out other
queen cells and sting the infant queens within. It is also thought
that, should two queens emerge simultaneously, they will fight to
the death. Recent studies, however, have indicated that as many
as 10% of Apis mellifera colonies may maintain two queens. The mechanism
by which this occurs is not yet known, but it has been reported
to occur more frequently in some South African subspecies of Apis
mellifera.[citation needed] Regardless, the queen asserts her control
over the worker bees through the release of a complex suite of pheromones
called queen scent. The queen lays all the eggs in a healthy colony. The number and pace of egg-laying is controlled by weather, availability of resources and the characteristics of the specific race of honey bee. Honey bee queens generally begin to slow egg-laying in the early fall and may even stop during the winter. Egg-laying will generally resume in late winter as soon as the days begin to get longer and peak in the spring. At the height of the season, the queen may lay over 2500 eggs per day – more than her own body mass. The queen fertilizes each egg as it is being laid into worker size cell using stored sperm from the spermatheca. Eggs, laid into drone size (larger) cells are left unfertilized. The unfertilized eggs have only half as many genes as the queen or worker eggs and develop into drones. Genome The
genome is unusual in having very few transposons, while they have
been present in the evolutionary past (inactive remains were found)
and in general evolved slower than in Diptera species. Pheromones Communication Another signal for communication is the shaking signal, also known as the jerking dance, vibration dance, or vibration signal. It is a modulatory communication signal because it appears to manipulate the overall arousal or activity of behaviors. The shaking signal is most common in worker communication, but it is also evident in reproductive swarming. A worker bee vibrates its body dorsoventrally while holding another honey bee with its front legs. Jacobus Biesmeijer examined the incidence of shaking signals in a forager’s life and the conditions that led to its performance to investigate why the shaking signal is used in communication for food sources. Biesmeijer found that the experienced foragers executed 92.1% of the observed shaking signals. He also observed that 64% of the shaking signals were executed by experienced foragers after they had discovered a food source. About 71% of the shaking signal sessions occurred after the first five foraging success within one day. Then other communication signals, such as the waggle dance, were performed more often after the first five successes. Biesmeijer proved that most shakers are foragers and that the shaking signal is most often executed by foraging bees over pre-foraging bees. Beismeijer concluded that the shaking signal presents the overall message of transfer work for various activities or activity levels. Sometimes the signal serves to increase activity, when bees shake inactive bees. At other times, the signal serves as an inhibitory mechanism such as the shaking signal at the end of the day. However, the shaking signal is preferentially directed towards inactive bees. All three types of communication between honey bees are effective in their jobs with regards to foraging and task managing. "The general story of the communication of the distance, the situation, and the direction of a food source by the dances of the returning (honey bee) worker bee on the vertical comb of the hive, has been known in general outline from the work of Karl von Frisch in the middle 1950s." Beekeeping Honey bees collect flower nectar and convert it to honey which is stored in their hives. The nectar is transported in the stomach of the bees, and is converted to honey through the addition of various digestive enzymes, and by being stored in a "honey cell" and then partially dehydrated. Nectar and honey provide the energy for the bees' flight muscles and for heating the hive during the winter period. Honey bees also collect pollen which supplies protein and fat for bee brood to grow. Centuries of selective breeding by humans have created honey bees that produce far more honey than the colony needs. Beekeepers, also known as "apiarists," harvest the honey. Beekeepers often provide a place for the colony to live and to store honey. There are seven basic types of beehive: skeps, Langstroth hives, top-bar hives, box hives, log gums, D.E. hives and miller hives. All U.S. states require beekeepers to use movable frames to allow bee inspectors to check the brood for disease. This allows beekeepers to keep the Langstroth, top-bar, and D.E. hives freely, but other types of hives require special permitting, such as for museum use. The type of beehive used significantly impacts colony health and wax and honey production. Modern hives also enable beekeepers to transport bees, moving from field to field as the crop needs pollinating and allowing the beekeeper to charge for the pollination services they provide. In cold climates some beekeepers have kept colonies alive (with varying success) by moving them indoors for winter. While this can protect the colonies from extremes of temperature and make winter care and feeding more convenient for the beekeeper, it can increase the risk of dysentery (see the Nosema section of diseases of the honey bee) and can create an excessive buildup of carbon dioxide from the respiration of the bees. Recently, inside wintering has been refined by Canadian beekeepers, who build large barns just for wintering bees. Automated ventilation systems assist in the control of carbon dioxide build-up. Products As an example, in California, the pollination of almonds occurs in February, early in the growing season, before local hives have built up their populations. Almond orchards require two hives per acre (2,000 m² per hive) for maximum yield and so the pollination is highly dependent upon the importation of hives from warmer climates. Almond pollination, which occurs in February and March, is the largest managed pollination event in the world, requiring more than one third of all the managed honey bees in the United States. Massive movement of honey bee are also made for apples in New York, Michigan, and Washington. And despite the inefficiency of honey bees in pollinating blueberries,[9] huge numbers are also moved to Maine for blueberries, because they are the only pollinators that can be relatively easily moved and concentrated for this and other monoculture crops. Commercial beekeepers plan their movements and their wintering locations with prime reference to the pollination services they plan to perform. Bees, as well as some other insects, are particularly beneficial as pollinators to most plants, as they maintain flower constancy, which means they are more likely to transfer pollen to other conspecific plants.[10] Also, flower constancy prevents the loss of pollen during interspecific flights and pollinators from clogging stigmas with pollen of other flower species. Honey Beeswax Pollen Propolis Royal
Jelly Hazards
and Survival
As an invasive species, feral honey bees have become a significant environmental problem in places where they are not native. Imported bees may compete with and displace native bees and birds, and may also promote the reproduction of invasive plants that native pollinators do not visit. Also, unlike native bees, they do not properly extract or transfer pollen from plants with poricidal anthers (anthers that only release pollen through tiny apical pores), as this requires buzz pollination, a behavior which honey bees rarely exhibit. For example, honey bees reduce fruiting in Melastoma affine (a plant with poricidal anthers) by robbing stigmas of previously deposited pollen. |

 Pollination
Pollination Environmental
Hazards
Environmental
Hazards|
|
||
|
|
||
|
|
||
|
|
||
| Canku Ota is a free Newsletter celebrating Native America, its traditions and accomplishments . We do not provide subscriber or visitor names to anyone. Some articles presented in Canku Ota may contain copyright material. We have received appropriate permissions for republishing any articles. Material appearing here is distributed without profit or monetary gain to those who have expressed an interest. This is in accordance with Title 17 U.S.C. Section 107. | ||
|
Canku Ota is a copyright ©
2000, 2001, 2002, 2003, 2004, 2005, 2006, 2007, 2008, 2009, 2010,
2011 of Vicki Barry and Paul Barry.
|
||
| |
|
|
|
The "Canku
Ota - A Newsletter Celebrating Native America" web site and
its design is the
|
||
|
Copyright ©
1999, 2000, 2001, 2002, 2003, 2004, 2005,
2006, 2007, 2008, 2009, 2010, 2011
of Paul C. Barry.
|
||
|
All Rights Reserved.
|
||